
我们的记忆取决于我们回忆关于世界的细节的能力——孩子的脸庞、一只鹅、一个湖泊。然而,为了将它们转化为真实的体验,大脑必须以某种方式将这些独立的元素融合成一个完整的整体——当她看到一群鹅突然从湖畔的芦苇丛中飞起时,孩子脸上的表情。
连贯的记忆感也依赖于其他因素。我们数千年的生存依赖于不仅回忆起正确的信息——比如,狮子或蛇——还要回忆起背景。我们是在非洲稀树草原的孤立地带遭遇了一场突如其来的动物遭遇战,还是在圣地亚哥动物园悠闲地观看?
为了避开我们日常生活中的其他类型的捕食者,我们也需要能够随着时间的推移连接记忆:判断一项看似有吸引力的投资是否值得追求,取决于推荐来源——例如,建议者的可靠性。未能将两者联系起来可能会导致灾难性的后果。
支持科学新闻报道
如果您喜欢这篇文章,请考虑通过以下方式支持我们屡获殊荣的新闻报道 订阅。通过购买订阅,您正在帮助确保有关塑造我们当今世界的发现和思想的有影响力的故事的未来。
神经科学领域开始努力解决大脑如何在空间和时间上连接记忆。到目前为止,绝大多数研究都集中在我们获取、存储、回忆和改变单个记忆的方式上。然而,大多数记忆并非作为单一、孤立的实体而独立存在。相反,一个回忆会召唤下一个回忆,建立起复杂的记忆序列,帮助我们更好地预测和理解我们周围的世界。
大脑用来创造这些关联记忆的基本机制正开始显现出来——这得益于我的实验室和其他实验室 20 年的研究。理解交织个体记忆所涉及的物理过程,不仅可以深入了解大脑的工作方式,还可以帮助我们预防破坏我们创造和联系记忆能力的记忆障碍。
意外的惊喜
当我们在 20 世纪 90 年代后期开始研究记忆关联时,我们缺乏解决这个问题所需的工具和基本知识。确定记忆如何相互交织的关键第一步是我们发现了一个称为记忆分配的概念,即大脑使用特定规则将学习信息的片段分配给大脑区域中参与形成记忆的离散神经元群体的认识。
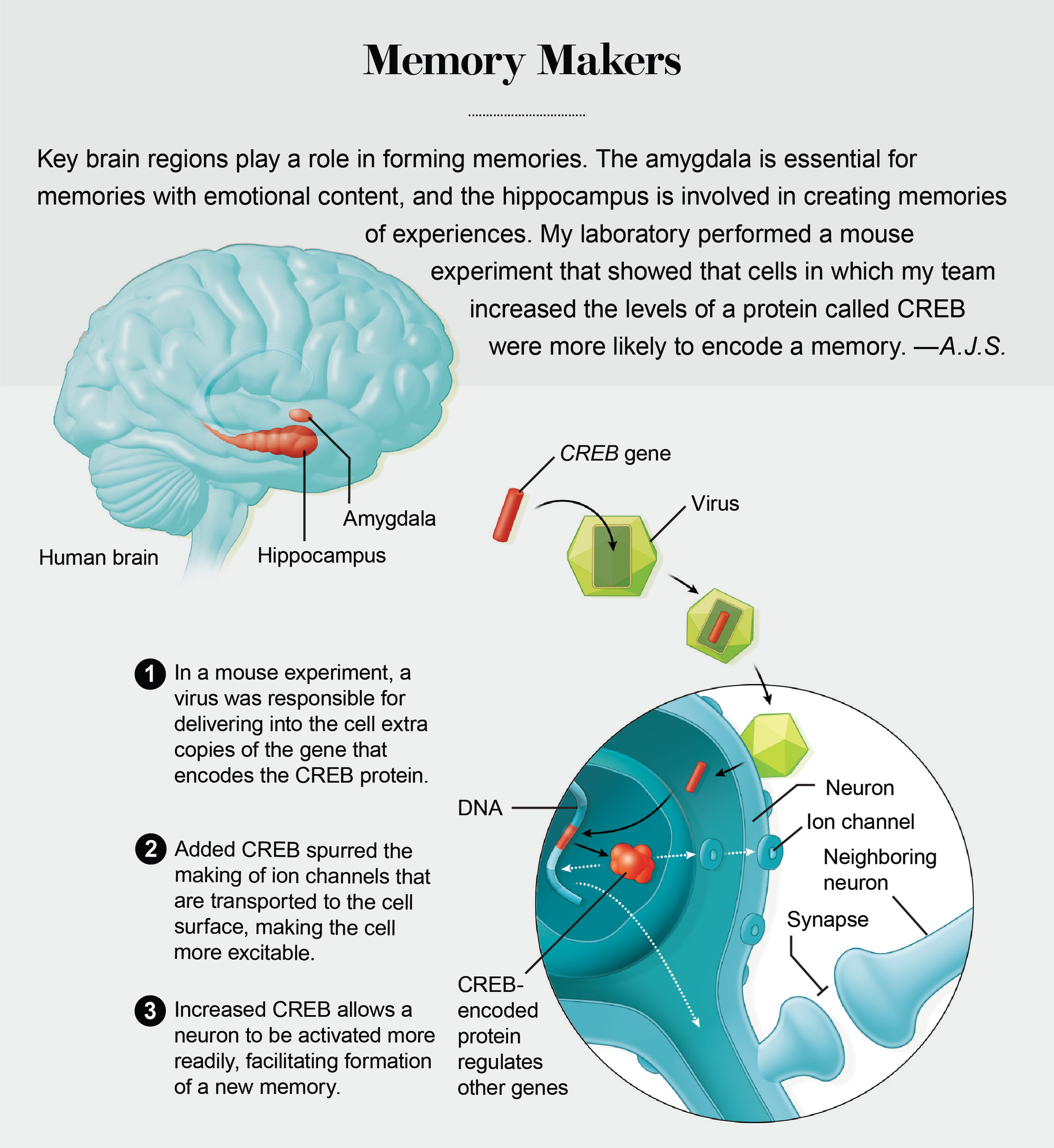
巧合在记忆分配的发现中发挥了关键作用。这始于 1998 年我在耶鲁大学访问期间与朋友兼同事、现任埃默里大学的迈克尔·戴维斯的一次谈话。戴维斯与我分享了他的实验室研究的发现,其中他的实验室操纵了一种称为CREB的基因,以增强大鼠的情绪记忆——例如,音调与电击之间的关联。此前,我的实验室(现在位于加州大学洛杉矶分校)和其他研究人员已经表明,形成长期记忆需要CREB基因。CREB 基因通过编码一种调节记忆所需其他基因表达的蛋白质来完成这项任务。在学习过程中,一些突触(神经元用来交流的细胞结构)会被建立或加强,以便它们可以促进细胞之间的相互作用。CREB 蛋白质充当这个过程的分子架构师。没有它的帮助,大多数经历都会被遗忘。
令我惊讶的是,戴维斯的研究小组能够改善记忆,即使他的实验室仅在杏仁核(大脑中对情绪记忆至关重要的区域)的整体神经元群体的一个子集中提高了 CREB 水平。在我访问耶鲁大学后的几个月里,一直困扰我的问题是,记忆是如何最终进入到少数可以利用更高 CREB 水平的细胞中的?难道 CREB 不仅协调了记忆的形成,还有助于确保具有 CREB 的细胞更有可能参与记忆的形成吗?在我们自己对 CREB 的研究中,我们将重点放在了我们已知与记忆相关的特定大脑区域内的功能:杏仁核和海马体。后者存储了一个人周围环境的内部地图。

显微镜安装在活体小鼠的头部,让研究人员检查存储记忆的大脑细胞的活动。图片来源:Denise J. Cai 加州大学洛杉矶分校整合学习与记忆中心
科学就像寻找问题一样,也像回答问题一样。与戴维斯的谈话帮助我意识到,神经科学家对给定记忆如何分配到处理和存储我们回忆的大脑区域的神经元中的规则知之甚少,如果真的有规则的话。因此,我们决定更仔细地观察。
我们的第一个重大突破是在我们招募了神经科学家希娜·A·乔斯林之后,她曾在戴维斯实验室研究 CREB。在她在我的实验室以及后来在多伦多大学的她自己的实验室与同事进行的一系列动物实验中,乔斯林使用病毒将 CREB 的额外拷贝引入小鼠杏仁核内的特定神经元。她表明,与邻近的神经元相比,这些神经元存储恐惧记忆的可能性几乎高出四倍。
2007 年,经过近十年的努力,我的实验室与乔斯林的团队合作,最终发表了证据,表明情绪记忆并非随机分配到杏仁核内的神经元。相反,被选来存储这些记忆的细胞是那些具有更多 CREB 蛋白质的细胞。同样重要的是,随后的实验表明,CREB 在其他大脑区域(包括海马体和大脑皮层,即最外层)也具有类似的功能。
开启和关闭记忆
为了证实 CREB 在记忆分配中的作用,我们转向了近年来改变了记忆研究的新开发的方法。这些实验室技术使得激活或关闭神经元成为可能——实际上,可以引发或沉默记忆。
举一个例子,当时在我的实验室工作的周宇对一小部分小鼠杏仁核神经元进行了基因改造,使它们具有更高的 CREB 水平,并表达了由加利福尼亚州拉霍亚市索尔克生物研究所爱德华·卡拉威实验室设计的另一种蛋白质。卡拉威的精巧蛋白质使我们能够在我们选择的时间沉默 CREB 神经元。当我们关闭具有高 CREB 的神经元时,留下蛋白质水平较低的对应神经元仍然活跃,情绪记忆被抑制,这一结果提供了证据,表明具有较高 CREB 水平的神经元更有可能参与记忆存储。
我们知道较高水平的 CREB 可以决定哪些细胞存储给定的记忆,但我们不知道这是如何发生的。斯坦福大学的罗伯特·马伦卡和他的同事发现,增加某些神经元中的 CREB 意味着它们更容易被激活。这种兴奋性的增加是否是具有较高 CREB 水平的神经元被选用于记忆存储的原因?
为了解决这个问题,周修改了杏仁核神经元以产生更多 CREB。使用微小的微电极,她确定了这些神经元有多容易被激活,这是兴奋性的一种度量。结果证实,与未改变的对应神经元相比,修改后的神经元更容易被开启。兴奋性的升高(增强了接收和传递在神经元之间传递信息的电脉冲的准备状态)表明,细胞可能更好地准备好启动记忆形成所需的一系列过程。
为了检验这个想法,周还研究了涉及具有更多 CREB 的神经元的突触连接。大量证据表明,突触连接强度的增加对于记忆形成至关重要。在对小鼠进行一项随后引发情绪记忆的任务训练后,她测试了杏仁核神经元的突触连接强度,这些神经元具有更高的 CREB 水平,看看它们是否比未被改变以产生更多 CREB 的细胞具有更强的连接。
为了做到这一点,她用微小的电流刺激这些细胞的突触,并用嵌入细胞内的微小电极记录它们的反应。正如预期的那样,具有较高 CREB 的杏仁核神经元比其他细胞具有更强的突触,这一结果与它们更有可能存储情绪记忆的想法一致。
在更新近的工作中,乔斯林的实验室证明,通过对一组预先确定的杏仁核神经元进行基因工程改造,使其具有一种特定类型的离子通道,可以增加这些神经元的兴奋性,从而将恐惧体验的记忆存储在其中。离子通道在细胞表面形成孔隙,乔斯林选择的特定离子通道允许这些细胞更容易被激活。同样,霍华德·休斯医学研究所珍妮莉亚研究园区(位于弗吉尼亚州阿什本)的神经科学家阿尔伯特·李的实验室报告说,当动物在跑道上跑动时,人为地增加特定位置的海马神经元的兴奋性,使这些神经元更有可能对跑道上的那个位置做出反应,这一结果与我们的发现一致,即兴奋性在决定哪些细胞参与存储给定记忆中起着至关重要的作用。
最后,我们的小组以及乔斯林的小组都利用了一项突破性技术,称为光遗传学,该技术使用光来激活或抑制神经元。我们使用该技术来开启具有较高 CREB 水平的特定神经元。托马斯·罗杰森和巴拉吉·贾亚普拉卡什,当时都在我的实验室工作,他们首先对杏仁核神经元进行基因工程改造,以产生更多 CREB 和视紫红质通道蛋白 2 (ChR2),这是一种由蓝光激活的离子通道。然后我们表明,当我们使用光开启具有较高 CREB 的杏仁核神经元时,我们可以人为地触发小鼠对恐惧记忆的回忆,但当开启蛋白质水平较低的神经元时则不行,这证实了记忆存储在这些神经元中。
连接起来
2009 年,我被要求写一篇关于我们记忆研究的文章,我借此机会介绍了我们关于记忆如何随时间推移而联系起来的想法。CREB 调节哪些细胞形成给定记忆的能力——换句话说,记忆分配——使我提出了一个假设,即这个过程可能是连接不同记忆的关键,我的实验室现在称之为“分配以连接”假说。由于记忆分配发生在具有更高 CREB 且更容易激活的神经元子集中,因此这个过程为这些神经元准备好存储另一个记忆。当两个记忆共享许多相同的神经元时,它们就会正式连接起来。
因此,在回忆两个记忆中的一个时激活这些神经元会触发对另一个记忆的回忆。这个想法的关键预测是,在一天之内形成的、时间上更接近的两个记忆比间隔较长时间的记忆更有可能被连接起来。如果间隔远大于一天,第二个记忆不再受益于第一个记忆触发的兴奋性,因此存储在不同的神经元群体中。记忆连接的时间限制性质是有道理的,因为一天之内发生的事件比相隔一周的事件更有可能彼此相关。
撰写文章和概述这些想法让我更加投入到我们如何检验它们的挑战中。“分配以连接”假说很简单,但我们如何证实其合法性根本不清楚。测试不得不等待合适的时机。
当丹尼斯·J·蔡和贾斯汀·肖伯(当时都在我的实验室工作)加入该项目时,情况发生了变化。蔡提出了一个聪明的想法。她与肖伯一起,在同一天内、在五个小时的间隔内,将小鼠暴露在两个房间中,希望这两个房间的记忆能够联系起来。后来,她在第二个房间里给了它们轻微的爪子电击。正如预期的那样,当她随后将小鼠放入它们受到电击的房间时,它们僵住了,大概是因为它们记得在那里受到了电击。小鼠会僵住作为对恐惧的自然反应,因为大多数捕食者在猎物移动时更容易注意到它们。
当蔡和肖伯将小鼠放入中性房间时,关键结果出现了。我们推断,如果两个房间的记忆联系在一起,中性空间中的小鼠会想起在另一个房间里受到电击,因此会预感到恐惧而僵住——这正是我们发现的。
我们还猜测,如果两个记忆间隔七天,它们就不太可能被连接起来。事实上,在较长时间间隔后,再次将动物暴露在中性房间中并没有让它们想起电击室,它们也没有僵住。一般来说,对于远大于一天的时间间隔,记忆仍然是未连接的。值得注意的是,加州大学圣地亚哥分校吉尔·鲁特格布实验室的研究表明,我们实验中使用的脑区非常能够区分我们在测试的时间框架内(数小时)的时间间隔,这一结果与我们的发现相符。
这些行为发现令人兴奋,但它们并没有检验该假说的关键预测——在紧密间隔形成的不同记忆存储在同一大脑区域的重叠神经元群体中。这种物理重叠将两个记忆联系起来,因此对一个记忆的回忆会让人想起另一个记忆。
可视化记忆
要真正检验“分配以连接”假说,就需要在大脑中看到记忆被创造出来的过程。用于对活体小鼠的神经元进行成像的技术已经在使用中,但它们都需要将小鼠的头部固定在大型显微镜上,这种设置不利于检验该假说所需的行为实验。
不过,我发现令人惊奇的是,在我的职业生涯中,正确的技术经常在我们最需要它的时候出现。我碰巧参加了加州大学洛杉矶分校举办的研讨会,斯坦福大学的马克·施尼策在研讨会上介绍了他实验室刚刚发明的一种微型显微镜,它可以可视化自由活动小鼠的神经元活动。这种两到三克重的显微镜可以像帽子一样安装在动物的头上。该仪器正是我们小组追踪给定记忆激活的神经元所需的。施尼策和他的同事已经表明,它可以用来跨时间追踪我们正在研究的神经元。因此,我们可以使用他们革命性的微型显微镜来检验我们的“分配以连接”假说的关键预测:在一个记忆期间活跃的许多神经元在几个小时后创建另一个连接记忆期间再次活跃起来。
我们对这项奇妙发明的承诺感到非常兴奋,因此我们决定设计我们自己的显微镜版本。我们与加州大学洛杉矶分校的佩曼·戈尔沙尼和巴尔吉特·卡克的实验室合作,共同聘请了一位才华横溢的博士后研究员丹尼尔·阿哈罗尼,他继续设计了我们后来称之为加州大学洛杉矶分校迷你显微镜的东西。与施尼策显微镜类似,我们的迷你显微镜配备了一个镜头,可以嵌入在我们想要记录的脑细胞附近。该设备卡在固定在动物头骨上的底板上,在训练任务和记忆测试期间保持稳定。正如我们从其他研究人员那里借用技术一样,我们也乐于分享。我们是科学界开源运动的积极支持者,并将我们用于加州大学洛杉矶分校迷你显微镜的设计和软件提供给了全球数百个其他研究小组。
为了可视化迷你显微镜的神经元活动,蔡和她的同事特里斯坦·舒曼利用了一种成像技术,该技术对动物的神经元进行基因工程改造,使其在细胞中的钙水平升高时发出荧光——这被称为基因编码钙指示剂。
我们决定专注于海马体的 CA1 区域,因为它在学习和记忆地点(例如我们在行为实验中使用的房间)方面发挥着作用。戴着迷你显微镜帽子的小鼠被放置在两个房间中。我们想知道暴露于不同房间之间的时间间隔是否会影响哪些神经元被激活。
图片来源:塔米·托尔帕
结果超出了我们的预期!基本上,我们的迷你显微镜和行为实验表明,当小鼠连接两个房间的记忆时,当动物访问第一个房间时变得活跃的许多 CA1 神经元在它们在第二个房间探索时也被开启。如果访问之间的间隔约为五个小时,小鼠会在相似的神经元簇中形成两个记忆。当时间间隔增加到七天时,这种重叠的激活模式没有出现。
我们对这个发现感到高兴,因为它证实了“分配以连接”假说的基本前提:当记忆存储在重叠的神经元群体中时,它们会耦合。如果您稍后重新激活为两个记忆中的任何一个形成的神经元集合,它会刺激另一个记忆并促进其回忆。
标记记忆
为了进一步验证迷你显微镜的结果,蔡转向了神经科学家马克·梅福德(现任斯克里普斯研究所)开发的另一种方法。这项实验涉及梅福德的技术,称为 TetTag 系统(代表四环素标记)。当转基因小鼠访问房间时形成记忆时,TetTag 会用荧光标记标记激活的神经元,该标记可保持完整数周。
然后,对动物的尸检研究可以比较最近激活的神经元——用记忆形成后立即表达的基因标记——与用长期标记标记的神经元。此步骤不仅识别出由一个事件开启的神经元——在这种情况下,一个神经元具有单个荧光标记——还识别出由两个事件激活的神经元:两个标记都发出荧光。
蔡和她的团队使用与以前相同的实验设置表明,在短短的五个小时间隔内,每个具有双重标记的记忆的编码神经元之间的重叠明显大于偶然预期的重叠。对于七天的时间间隔,两次体验之间的重叠没有明显高于偶然水平。
乔斯林的加拿大多伦多研究小组进行的其他实验提供了更多证据来证明我们的记忆连接假说的有效性。她的研究小组不仅进行了不同版本的神经元标记实验,科学家们还发现了记忆连接的独立行为证据。多伦多研究人员推断,如果编码两个记忆的神经元群体重叠,第一个记忆触发的 CREB 水平的增加也会加强第二个记忆。但是,乔斯林的团队没有像我们的工作那样将小鼠暴露于不同的地点,而是训练动物学习识别两种不同的音调。如果两个训练课程发生在六小时内,而不是六到二十四小时内,则对第一个音调的训练会加强对第二个音调的记忆。
最近,日本富山大学的猪口薰和他的同事们将这项分析更进一步。他们使用光遗传学来灭活两个不同情绪记忆共享的细胞群,同时保持其他细胞不受干扰,包括那些对每个记忆都是唯一的细胞。研究人员表明,通过灭活共享细胞,他们能够破坏两个记忆之间的连接,而不会影响对每个个体记忆的回忆。这项优雅的实验直接证明了两个记忆共享的神经元是记忆连接的关键。它也增加了为我们初步的“分配以连接”假说提供独立证据的实验室数量。
改善衰老过程中的记忆
接下来,我们决定研究老年小鼠的记忆连接。与年轻小鼠相比,老年小鼠大脑(包括海马体 CA1 区域的神经元)中的 CREB 水平较低,因此兴奋性也较低。了解了这一点,我们预测老年小鼠在连接记忆方面应该会遇到困难。因此,蔡和她的同事着手重复我们在老年动物身上已经完成的许多相同实验。结果让我们感到惊讶。经验丰富的科学家知道,假说只是工具。我们不期望它们一定是正确的。不可避免的失败帮助我们沿途重塑我们的想法。但这一次,我们的预感被证明是正确的。
.png?w=2000)
图片来源:塔米·托尔帕(插图);来源:“共享神经元集合连接时间上接近的不同情境记忆”,丹尼斯·J·蔡等,《自然》,第 534 卷;2016 年 6 月 2 日(神经元图像)
我仍然记得蔡气喘吁吁地冲进我的办公室。她告诉我,中年小鼠尽管记得每个房间,但确实在连接记忆方面存在问题,即使它们在相隔五小时的时间内暴露于房间中,这个间隔对于年轻小鼠来说没有困难。与年轻成年小鼠相比,老年动物的迷你显微镜成像显示存储记忆之间缺乏重叠。
我们既兴奋又持怀疑态度,因此我们立即回去重复了实验。第二次,结果变得更加令人信服。CREB 水平较低的中年小鼠的神经元不如年轻小鼠的神经元容易连接记忆。
这些结果鼓舞我们扩大研究范围。我们能否在老年小鼠探索两个房间时,人为地增加 CA1 神经元子集的兴奋性,确保在一个房间中激活的一些 CA1 神经元在动物移动到第二个房间时也被开启?
为了实现这一点,我们利用了一种突破性的方法,该方法在细胞表面对受体进行基因工程改造,从而可以控制细胞的功能。该技术有一个令人难忘的技术缩写词 DREADD(代表设计师受体专门被设计师药物激活)。激活 DREADD 受体使我们能够在动物探索两个房间时开启同一组 CA1 神经元,从而在它们对两个围栏的记忆之间建立联系。
我必须承认,起初这个实验的想法听起来很荒谬。有很多原因可能导致它失败。首先,对地点的记忆涉及数百万个神经元,这些神经元分布在多个相互连接的大脑区域,而不仅仅是 CA1 区域。衰老可能会影响许多(如果不是全部)这些区域的记忆连接过程。因此,即使我们成功地增加了 CA1 神经元子集的兴奋性,这些细胞也可能不是正确的细胞。更重要的是,我们可能没有触发正确水平的兴奋性。
但实验成功了。这种“孤注一掷”试验的关键在于平衡时间和金钱的投入与可能带来的潜在回报。然而,在这种情况下,我可以肯定地说,幸运之神站在我们这边。通过恢复中年小鼠特定 CA1 神经元子集中兴奋性的增加,我们能够将两个记忆分配给许多相同的 CA1 神经元,从而恢复这些中年小鼠的记忆连接。
来自啮齿动物和人类的其他实验室的研究也阐明了一个记忆如何与另一个记忆交织在一起。波士顿大学已故神经科学家霍华德·艾兴鲍姆证明,大鼠能够找到共享内容的记忆之间的联系。德克萨斯大学奥斯汀分校的神经科学家艾莉森·普雷斯顿和她的同事表明,当记忆共享内容时,人类可以更容易地将它们联系起来。回忆起一个可能会带回另一个。2017 年,同一研究小组还表明,时间上的接近性促进了跨时间的记忆整合,这一结果在人类身上与我们在小鼠身上的结果惊人地相似。
我们可支配的用于测量和控制神经活动的工具库不断增长,这开始揭示我们大脑用来组织信息的机制。我们的团队现在正试图以新的方式扩展这项工作。我们正在与希腊研究与技术基金会分子生物学与生物技术研究所的计算神经科学家帕纳约塔·波拉齐合作,构建计算机模型来模拟记忆如何以及何时连接起来。我们也在试图弄清楚控制不同大脑结构中记忆连接所需时间间隔的机制。
到目前为止,多个实验室进行的大量广泛实验都强烈支持“分配以连接”假说。我们希望对记忆如何纠缠在一起的理解可能有助于我们开发治疗记忆问题的疗法,这些记忆问题在广泛的精神疾病中都很常见,从与年龄相关的认知衰退到精神分裂症、抑郁症和双相情感障碍。除了临床意义之外,我们描述的研究还反映了记忆研究中一个激动人心的新时代,在这个时代,我们所做的实验不再受我们拥有的技术的限制,而是受我们想象力的范围的限制。