
一只孤独的霸王龙 嗅着白垩纪潮湿的空气,嗅到了树林那边一群三角龙正在吃草的气味。当这只掠食者扫描洪泛区时,它的视力突然聚焦。一只三角龙脱离了兽群,游荡到了袭击范围之内。霸王龙静静地站着,制定了一个攻击计划,预测它必须以精确的角度与目标相交,才能在三角龙重新回到兽群的安全地带之前发起攻击。下午的寂静被打破了,掠食者冲破森林边缘的低矮树枝,开始了猛烈的追击。
霸王龙在如此多的书籍、游戏和电影中猎杀三角龙,以至于这种遭遇已经变得老生常谈。但是,这样的场景真的在现实生活中发生过吗?霸王龙会通过视觉还是嗅觉来识别猎物?三角龙会被响亮的树枝断裂声警告,还是因为无法定位声音来源而依然浑然不觉?霸王龙会像猫一样计划攻击,还是会像鲨鱼一样不分青红皂白地猛扑过去?
自从恐龙在19世纪早期首次被描述以来,古生物学家一直在争论它们的智力、感官能力和行为复杂性。早期的研究依赖于天然颅腔内模,这是当沉积物填充头骨中的空隙时形成的铸型。这些铸型复制了生物体内的颅腔内容物的形状。长期以来的传统观点认为,所有恐龙的大脑都很小,因此行为也很简单。关于恐龙智力最有趣的例子可能来自19世纪的古生物学家奥瑟内尔·查尔斯·马什,他假设装甲恐龙剑龙在臀部附近有一个第二个大脑,以补充其头骨中核桃大小的大脑。这个想法是基于恐龙骨盆附近脊髓管的模糊的颅腔形状的扩张。这种神秘的扩张现在被认为是糖原体——一种储存富含能量的葡萄糖的结构,在一些现代鸟类中也存在于相似的位置。
支持科学新闻报道
如果您喜欢这篇文章,请考虑支持我们屡获殊荣的新闻报道,方式是 订阅。通过购买订阅,您正在帮助确保未来能够继续报道关于塑造我们今天世界的发现和想法的具有影响力的故事。
当今的古生物学家仍然不相信剑龙有能力进行更高层次的推理。但在最近几年,科学家们对其他一些恐龙,特别是那些产生鸟类的兽脚亚目谱系的成员的认知能力的评价有所提高。随着微型计算机断层扫描 (CT) 扫描等新技术的出现,我们现在可以重建大脑的体积和表面地形,而不必完全依赖于稀有的天然颅腔内模,这大大扩展了可供研究的物种数量。先进的成像技术还在教我们恐龙可能如何使用它们的大脑。我们现在拥有了回答问题的工具,即早已消失的动物如何感知周围的世界,以及在恐龙时代掠食者与猎物相遇时到底发生了什么。
霸王龙在迟钝的剑龙和会使用工具的乌鸦之间的智力谱系中处于什么位置?在去年秋天发表的一篇备受瞩目的论文中,范德比尔特大学的神经科学家苏珊娜·赫库拉诺-霍泽尔认为,霸王龙的智力大约相当于狒狒——这是一个令人震惊的结论,因为灵长类动物,以其庞大的大脑,是一些最聪明的动物。我们花了很长时间思考大脑体积如何随身体大小缩放,以及这种关系对于已灭绝的恐龙和鸟类的大脑功能意味着什么,我们很感兴趣地看到了关于这项研究的头条新闻。从表面上看,暴君蜥蜴王的大脑与它的身体大小相比显得相当渺小。这种六吨重恐龙的大脑重量不到一磅,与非洲象的11磅大脑相比显得微不足道,尽管非洲象是现存最大的陆地哺乳动物,但它的身体比霸王龙小。
赫库拉诺-霍泽尔认为,当涉及到智力时,大脑大小和身体大小之间的关系并不重要。她说,重要的是端脑中神经元的原始数量,端脑是大脑前部的一个区域,不仅包括处理气味的嗅球,还包括大脑皮层,更高层次的认知功能(如决策)发生在这里。科学家们之前对脊椎动物大脑中存在多少神经元只有粗略的了解,因为在不同的物种中,它们在大脑的不同部位的密度可能更高或更低。
拥有灵长类动物智力的霸王龙将是可怕的。然而,我们认为应该有一些注意事项。
赫库拉诺-霍泽尔和里约热内卢联邦大学的罗伯托·伦特发明了一种计算神经元的技术,称为各向同性分馏器法。它使用特殊的化学物质来溶解大脑,基本上是制作脑汤。一种荧光染料会给神经元的细胞核染色,使其发光并易于观察。研究人员可以精确地计算出汤中少量均匀样本中发光的细胞核,然后推断出活体大脑中神经元的总数。使用这种方法,赫库拉诺-霍泽尔和她的同事计算出人脑大约有1000亿个神经元,证实了早期的估计。
各向同性分馏器法很巧妙,但科学家们永远不会有实际的霸王龙大脑来溶解。相反,赫库拉诺-霍泽尔依赖于现存温血和冷血物种的端脑大小和神经元数量之间的比例关系,根据已灭绝恐龙的推断代谢率,将它们代入两个方程中的一个。这种比例关系在脊椎动物中差异很大。冷血或外温物种的神经元密度往往低于温血或内温物种。
例如,各向同性分馏器数据表明,一只159磅重的尼罗鳄大约有8150万个神经元,而一只73磅重的鸸鹋大约有13亿个神经元——几乎是鳄鱼的16倍,尽管只有它一半的大小。赫库拉诺-霍泽尔认为,大多数兽脚亚目恐龙可能是内温动物,因此接近神经元密度连续体的鸟类端。在这种假设下,一个重约12盎司的霸王龙端脑将包含大约30亿个神经元,与许多灵长类动物相当。

梅萨·舒马赫
拥有灵长类动物智力的霸王龙将是可怕的。然而,我们认为应该有一些注意事项。除了兽脚亚目恐龙是内温动物的合理推测之外,30亿个神经元的估计还依赖于其他一些假设。其中之一是整个颅腔都被大脑占据,这显然与霸王龙颅腔内模的形态不符。化石证据表明,在许多恐龙中,鼻窦和血管等结构占据了颅腔的很大一部分。实际的大脑体积将小于原始的颅腔体积。事实上,基于现代鳄鱼的研究表明,霸王龙的大脑可能只占整个颅腔体积的30%。
同样重要的是要注意,不同的脑区具有不同的功能。有些脑区致力于呼吸调节等基本任务,而另一些脑区则支持语言等复杂功能。如果大脑皮层在一个物种的大脑大小中占主导地位,而扩大的嗅球在另一个物种的大脑大小中占主导地位,那么具有相同端脑大小的两个物种可能具有截然不同的认知能力。对于现存物种,可以使用细胞染色或磁共振成像等技术来确定不同脑区之间的边界。
霸王龙真的能够嗅到风,并在看到猎物或腐肉之前很久就识别出它们,以便进行清扫。
对于恐龙来说,定义这些边界要困难得多,因为我们所能利用的只是表面拓扑结构。我们中的一位(巴拉诺夫)花费了她职业生涯的很大一部分时间来绘制骨性地标,这些地标可以更好地估计颅腔内模主要脑区的体积。这项工作表明,大脑皮层的扩张出现在更特化的兽脚亚目恐龙中,如窃蛋龙和驰龙——这些谱系的分支时间比暴龙科晚得多。相比之下,较早分化的兽脚亚目恐龙,如霸王龙,其大脑皮层相对较小,整个端脑的大部分都让给了嗅球。
一旦我们考虑到颅腔内模中容纳的非神经组织体积,霸王龙的端脑中不太可能有任何类型的30亿个神经元。我们同意霸王龙 是一个熟练的掠食者,但我们认为它可能不具备灵长类动物身上看到的先进的计划能力或协调的社会狩猎能力。
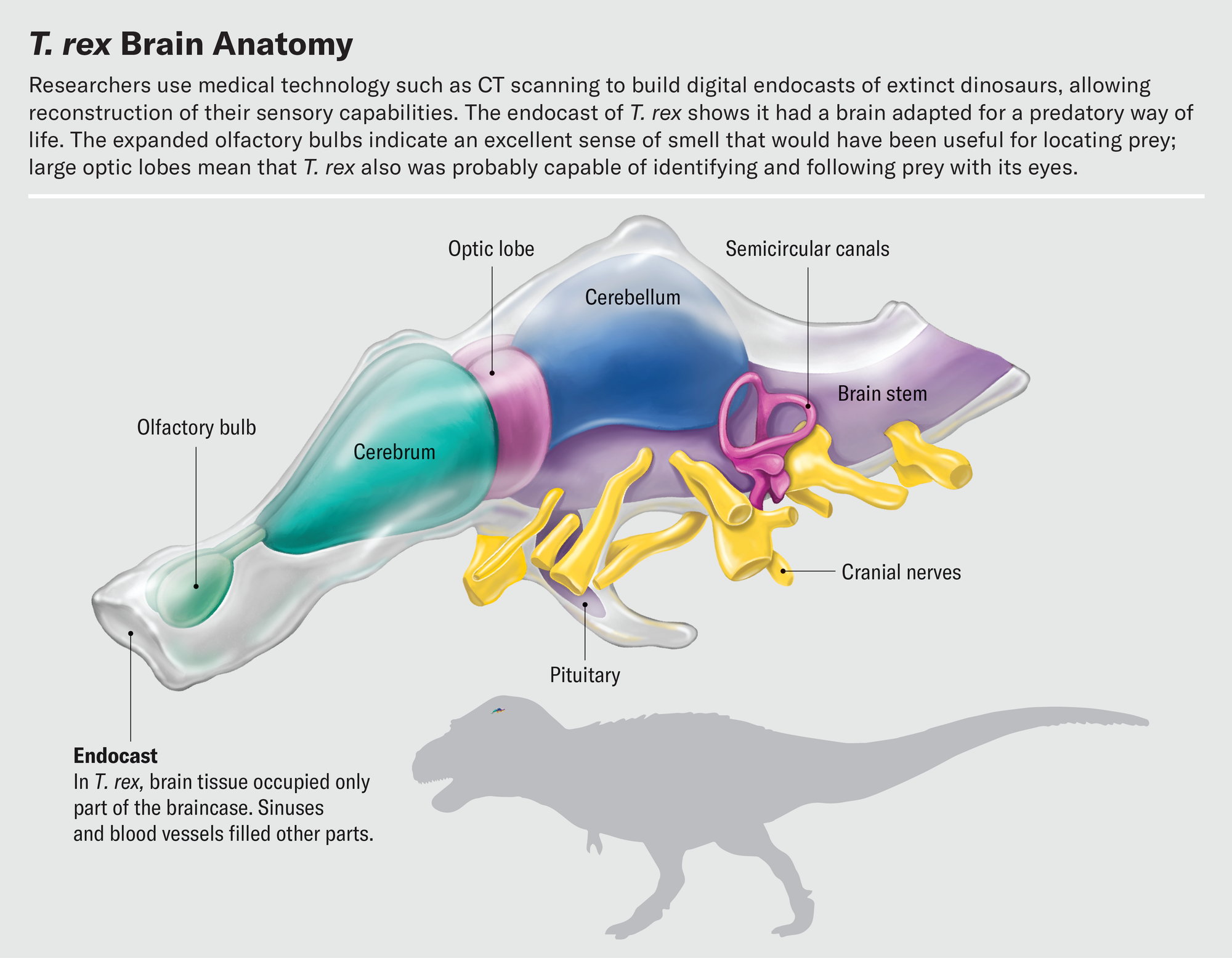
使用CT扫描构建的化石颅腔内模进行研究的最好之处之一是,我们可以研究内部特征,而不会损坏化石本身。虚拟地切割化石颅腔内模是一种度过下午的愉快方式。训练有素的古生物学家通过逐片探索大脑切片,可以使用骨性地标来解读关键脑区的边界,并以数字方式隔离这些区域。当我们从前到后分割大脑时,我们遇到的第一个结构是嗅球。嗅球的形状在恐龙及其亲属中差异很大。鳄鱼的嗅球大约有小葡萄大小,位于通往大脑其余部分的长柄末端。大多数鸟类的嗅球要小得多;事实上,在许多物种中,它们几乎无法与大脑皮层的其余部分区分开来。
顾名思义,嗅球促进嗅觉——一种依赖于称为气味分子的微小分子的感觉。吸入的气味分子与鼻组织中的受体结合,鼻组织通过神经元与嗅球进行通信。令人惊讶的是,每个受体都会产生一种气味分子-受体蛋白,这种蛋白会根据特定类型的气味分子进行调整。这些蛋白质中的每一种都由不同的嗅觉受体基因编码。基因组测序显示,鸟类拥有182到688个功能性嗅觉受体基因。
在最近的一项研究中,都柏林大学学院的格雷厄姆·休斯和约翰·菲纳雷利调查了恐龙的嗅觉。恐龙的嗅觉受体基因数量无法直接测量,但由于嗅球大小与受体数量相关,因此嗅球尺寸可以作为衡量动物探测气味能力的指标。休斯和菲纳雷利发现,总的来说,恐龙的嗅球比鸟类大得多。在兽脚亚目恐龙中,杂食性似鸟龙科动物的嗅球最小,而肉食性物种的嗅球最大。霸王龙名列前茅,其嗅球尺寸与存在600多个嗅觉受体基因的情况相符。这个数字与家猫相当,并且高于几乎所有现代鸟类。我们假设的跟踪三角龙的霸王龙真的能够嗅到风,并在看到猎物或腐肉之前很久就识别出它们,以便进行清扫。
梅萨·舒马赫;资料来源:劳伦斯·威特默和瑞安·里奇利在《解剖学记录》第292卷中发表的“关于暴龙(恐龙,兽脚亚目)大脑、颅腔和耳朵区域的新见解,及其对感官组织和行为的影响”,2009年;以及阿什利·莫哈特在俄亥俄大学2016年博士论文中发表的“粗略解剖脑区近似 (GABRA):评估已灭绝的初龙的大脑大小、结构和进化”(颅腔内模参考文献)
其他基于CT的颅腔内模研究暗示了霸王龙的眼睛有多么敏锐。通过虚拟地切割颅腔内模并隔离视叶,我们发现这些结构在已灭绝的兽脚亚目恐龙和现存鸟类中的相对大小相似。因此,鸟类一定是从它们的非鸟类祖先那里继承了它们的视觉敏锐度。众所周知,鸟类是高度视觉化的动物——一只鹰可以从半英里外发现一只兔子,一只飞行中的燕鸥可以追踪到水面下半英寸长的鱼,并从上方将其捕获。这种对视觉的依赖反映在鸟类大脑的结构中。视叶处理视觉信息并位于大脑皮层之后,是大脑最突出的特征之一。通常情况下,一个区域相对于大脑其余部分越大,该区域对动物就越重要。这对于视叶来说当然是成立的。
我们可以从已灭绝恐龙的进化关系中推断出它们的一些视觉能力。例如,鸟类和鳄鱼(已灭绝恐龙最近的现存亲戚)都具有看到颜色的视网膜受体类型。因此,恐龙很可能也具有色觉。然而,视觉是一种复杂的感觉。准确地重建已灭绝恐龙的视觉能力需要我们超越这些类型的推断。
使猛禽如此擅长捕捉猎物的一个因素是立体视觉——一种增强的感知深度的能力。这种复杂能力背后的感官适应令人惊讶地简单。它仅仅与眼球的位置有关。眼睛位于头部侧面的动物,如壁虎,缺乏重叠的视野,因此它们的三维视觉不好。眼睛位于头部前方的动物具有在鼻子前方重叠的视野。在这个重叠的空间内,每只眼睛从略微不同的角度感知相同的信息,产生所谓的双眼视觉。
尝试将目光固定在离您大约一英尺左右的物体上,然后闭上一只眼睛,再闭上另一只眼睛。物体似乎会移动,因为您在鼻子前面看到了相同的图像,但角度不同。大脑整合这些略有不同的图像以产生视觉深度。眼睛横向定位的动物通过用一只眼睛看某物,移动头部,然后用另一只眼睛看它来判断深度——这不是一种特别隐秘的技术。对于掠食者来说,双眼视觉特别有用,因为它允许动物识别并锁定猎物,而无需通过移动头部来暴露其位置。
为了确定已灭绝的动物是否具有立体视觉,我们必须考虑它们的眼眶位置,眼眶是头骨中容纳眼球的空间。俄勒冈大学的肯特·史蒂文斯对这个问题采取了一种创造性的方法,他根据几种兽脚亚目恐龙的骨骼结构雕刻了它们的头部。从那里,他能够绘制出它们的视野范围,包括任何障碍物,如角或特别大的鼻子。他发现霸王龙的眼睛朝前,鼻子狭窄,不会阻碍其视线,使其视觉敏锐度与鹰相似。与电影《侏罗纪公园》中虚构的霸王龙不同,真正的霸王龙不需要猎物移动才能将其从背景中挑出来。恐爪龙类,如伤齿龙和迅猛龙,可能更擅长识别猎物。由于它们的头部略微向前倾斜,它们拥有更宽广的视野和增强的深度感知能力,可能与猫头鹰媲美。
我们可以从已灭绝动物的眼眶中推断出比感知深度更多的信息。这些开口的大小及其相关的骨骼提供了关于它们所容纳的眼球大小的线索。较大的眼眶通常表明眼球较大。相对于头部大小而言,较大的眼球在夜行动物中很常见,因为它们可以容纳更多的视网膜感光细胞。此外,包括鱼类、一些已灭绝的哺乳动物和爬行动物(包括非鸟类恐龙和鸟类)在内的许多动物的眼球,在其被称为巩膜的纤维外层内嵌有骨性或软骨环。由于骨骼是坚硬的,这些巩膜环限制了眼球的运动,包括瞳孔可以张开的程度。这会影响到达视网膜感光细胞的光量。较大的巩膜环允许瞳孔张得更大,从而让更多的光线进入眼睛。
几年前,巴拉诺夫曾是南非威特沃特斯兰德大学的乔纳·乔伊尼尔领导的一个团队的成员,该团队研究了非鸟类恐龙眼眶中这些环的大小,以确定它们是喜欢在白天还是晚上活动。由于巩膜环不附着在头骨的任何部分,它们很容易与骨骼的其余部分分离,并且很少保存在化石中。在保留它们的罕见标本中,它们并不总是被发现处于原位。在这项研究中,研究人员使用了具有保存完好的巩膜环的已灭绝恐龙的CT图像,以数字方式隔离这些骨骼并在眼眶内重建它们。他们的结论是,它们的眼眶和巩膜环的比例表明,许多已灭绝的恐龙主要在白天活动。
但是,一种家猫大小的阿瓦雷兹龙科兽脚亚目恐龙,沙漠爪龙,提供了不同的结果。该团队检查了在蒙古晚白垩世沙漠沙丘沉积物中发现的一个沙漠爪龙头骨,并惊讶地发现其适应性与最著名的现存夜行动物之一——谷仓猫头鹰趋同。谷仓猫头鹰有大的眼眶和充足的巩膜环,这使得它们的瞳孔可以张得非常大。这种结构可以让大量的光线涌入眼睛,在最黑暗的条件下提供图像。在阿瓦雷兹龙科动物中也存在这些相同的特征表明,它们也是夜间活动的掠食者。

梅萨·舒马赫
没有人正式分析过霸王龙的眼眶形态,以深入了解这种恐龙最活跃的时间。然而,我们确实知道,霸王龙有一个很大的眼眶,形状几乎像一个钥匙孔。如果眼睛填满了整个眼眶,那么我们或许可以推断出霸王龙是夜间活动的,但来自近缘物种的巩膜环表明,它的眼球只填满了眼眶的一小部分——因此可能无法收集足够的光线在夜间发挥太大作用。事实上,英国伯明翰大学的斯特凡·劳特enschlager领导的头骨应力数字分析表明,霸王龙的大眼眶有助于分散其强力咬合产生的巨大应力,而不是容纳一个大眼睛。
在脊椎动物头骨的后部附近是一个有趣而复杂的结构:内耳。虽然从技术上讲它不是大脑的一部分,但它是一个重要的感觉器官,向大脑发送大量信息。两种特殊感觉,平衡和听觉,由内耳所谓迷路的不同部分控制。迷路包括半规管、环状结构,用于检测头部的旋转运动;前庭,一种球状结构,用于感知前后和左右运动;以及耳蜗,用于感知声音振动。内耳充满了液体,并利用该液体内毛细胞的偏转来检测这些不同类型的信息。
2021年,现任职于史密森尼学会的迈克尔·汉森和他的同事对半规管和前庭的形状进行了复杂的分析,以推断非鸟类恐龙的主要运动方式。他们制作了内耳空间的虚拟颅腔内模,以估计迷路在生命期间的形态。他们的数据表明,大多数恐龙的活动仅限于在地面上行走和奔跑。但是在导致鸟类的谱系中,耳朵的结构发生了变化。在其他变化中,半规管变长了。这种伸长允许恐龙做出和解读更复杂的头部运动。霸王龙没有长的半规管,这表明它只能行走或奔跑。但是,一些更接近迅猛龙的伤齿龙科恐龙的耳朵可以感知与飞行相关的复杂运动。这些恐龙可能能够滑翔或进行雏形的飞行,在现代鸟类起飞之前在空中移动。
我们可以从耳蜗中寻找恐龙听力的线索。耳蜗的长度与听觉灵敏度相关。较长的耳蜗管允许基底乳突的伸长,基底乳突是容纳拾取声音振动的毛细胞的结构。蜥蜴和乌龟往往具有短而粗壮的耳蜗,最适合检测低音调的声音。相比之下,鳄鱼和鸟类具有长而细长的耳蜗,擅长检测高音调的声音。
许多鸟类会唱出悦耳的歌曲来吸引配偶和保卫领地,因此它们对高音调发声的敏感性提高似乎是理所当然的。然而,耳蜗的伸长并非起源于鸟类,而是起源于鸟类和鳄鱼的共同祖先。奇怪的是,鳄鱼的发声仅限于低音调的闭口咕噜声——而不是细长的耳蜗擅长检测的那种声音。与其他爬行动物相比,鳄鱼为何具有如此先进的耳蜗形状一直是个谜。
汉森和他的合作者提出了一个巧妙的解释。他们假设,将鳄鱼、已灭绝的恐龙和鸟类与早期分化的爬行动物区分开来的耳蜗伸长与亲代抚育的进化有关。与大多数爬行动物不同,鳄鱼会照顾它们的幼崽。与大多数幼年爬行动物不同,幼年鳄鱼会发出唧唧声以引起父母的注意。也许鸟类和鳄鱼的祖先需要能够探测高音调的声音,以便听到它们的幼崽而不是配偶或竞争对手。
与食草的猎物相比,霸王龙的大脑比例较大,这与现代掠食者相似。
这个假设对鸟鸣的进化具有令人兴奋的意义。在这种情景中,幼年恐龙可能会发出唧唧声以引起注意,但成年恐龙的叫声仅限于简单的低音调叫声。在恐龙向鸟类过渡的过程中,一些谱系将高音调发声的能力保留到了成年期。耳蜗伸长最初因其在增强亲代抚育中的作用而受到自然选择的青睐,然后作为一种预适应,使歌曲能够在后来的鸟类中出现。
但是进化并非朝着单一方向发展。通过筛选恐龙数据,汉森和他的同事偶然发现了一种不寻常的短耳蜗,它存在于异特龙(霸王龙的近亲)中。这一发现表明,这种大型兽脚亚目恐龙失去了对高音调声音的敏感性。该团队推测,异特龙可能没有照顾它们的幼崽,从而消除了听到它们发声的选择压力。
耳蜗的形状也帮助我们了解了那些古怪的夜行性阿瓦雷兹龙科动物的生态。巴拉诺夫和她的同事发现,沙漠爪龙的耳蜗非常长,以至于它在头骨底部下方卷曲。已知夜行性鸟类(如猫头鹰)具有类似的结构。沙漠爪龙的过度伸长的耳蜗管表明,它的听力适应于非常高频的声音,例如昆虫发出的声音。该团队得出结论,这种小型恐龙在亚洲中部沙漠沙丘的黑暗中漫游,捕猎这些小型猎物。
人们倾向于将古生物学视为一门基于野外研究的学科,专注于在偏远沙漠地区度过夏天的浪漫魅力,手持鹤嘴锄,收集对科学界来说是全新的早已灭绝的动物化石。但是,如今,古生物学家很可能在实验室中使用生物医学和神经科学的前沿技术来进行他们最重要的发现。正是这些不同方法的结合使我们能够重建当霸王龙 遇到三角龙时可能真正发生的事情。
我们自己的研究告诉我们,与现代掠食者相似,霸王龙的大脑比例较大,这与食草的猎物相比。它大脑的很大一部分致力于嗅觉,因此霸王龙可能确实会嗅探空气以寻找它的下一顿饭,无论是沿着树林线吃草的活三角龙,还是已经死亡并在阳光下腐烂的三角龙。一旦霸王龙识别出气味,它就可以用它的立体视觉扫描地平线,寻找任何潜在猎物的迹象。它的眼睛将能够锁定那只毫不在意地在远离兽群安全地带的植被丛中觅食的三角龙。
当霸王龙冲过树林时,它可能会惊吓到一只在附近树枝上筑巢的小型伤齿龙科恐龙。凭借其扩大的内耳迷路提供的增强的运动技能,伤齿龙科恐龙可能会从巢穴中滑翔出来,转移掠食者对其唧唧叫的幼崽的注意力。三角龙可能因这种骚动而感到不安,停止了平静的吃草,并返回兽群的安全地带。这仍然是一个老生常谈的故事,但这是一个更具科学依据的故事。